

Dinosaur Cognition: Not Set in Stone
Kate Billow
Illustrations by Anna McFadden

Dinosaur Cognition: Not Set in Stone
Dinosaurs have been brought back to life. The prehistoric creatures roam free across a small strip of islands to which travel is strictly prohibited. The scientific disasters that transpired at the Jurassic Park and World theme parks have left Chris Pratt and Bryce Dallas Howard with an impossible task: to venture to one of the islands to save the dinosaurs from a volcanic eruption. This dangerous mission drives the plot of Jurassic World: Fallen Kingdom (2018), the fifth installment in the Jurassic Park film franchise, which has enraptured audiences since 1993 [1, 2]. A central challenge of the film is the cleverness of Chris Pratt’s favorite Velociraptor, Blue, who learns complex tasks and demonstrates emotional intelligence like a toothy, lethal golden retriever. Blue is touted as ‘potentially the second most intelligent piece of life on this planet,’ inflating the Velociraptor’s intelligence to near-human levels [2, 3]. While the Jurassic Park films suggest the possibility of highly intelligent dinosaurs, another widely held belief depicts them as mindless hunting machines mercilessly swept from the planet 66 million years ago [4, 5]. How have these two vastly different views formed about dinosaur intelligence? As the largest, most diverse, and most abundant creatures on the planet, dinosaurs dominated the planet for a remarkable 160 million years, and it may have been a considerable cognitive prowess that supported their longstanding reign [4, 6, 7]. However, fossil evidence does not indicate that dinosaurs ever achieved classic benchmarks of human intelligence, such as multi-component tool use, social group organization and cooperation, or complex language [8, 9]. So, how intelligent were the dinosaurs? The field of paleoneurology has emerged to tackle this question, relying on techniques similar to those of anthropologists, archaeologists, and neuroscientists to investigate dinosaurs’ cognitive capabilities [10]. Granted, these methods primarily come from fields focused on human life and inevitably color our understanding of the prehistoric creatures at the expense of the unique contexts in which they actually lived [5, 10,].
Genius Genus: Who Were the Dinosaurs?
The term ‘dinosaurs’ is a colloquial catch-all for the scaly creatures that ruled the planet long before humans, but describes a very diverse group of organisms [6, 11]. Dinosaurs were reptiles who lived from roughly 230 to 66 million years ago, when an asteroid struck the Yucatán Peninsula in modern-day Mexico, wiping out three-quarters of all animal and plant species [8, 12, 13]. Birds are direct descendants of the only dinosaurs to survive this extinction event, so your pet chicken or the hummingbird at your bird feeder is technically a living dinosaur [14, 15, 16]! Since we do not live in the fictional world of Jurassic Park and cannot directly evaluate dinosaurs, birds are useful proxies for inferring dinosaur brain anatomy and function [8, 10, 17].
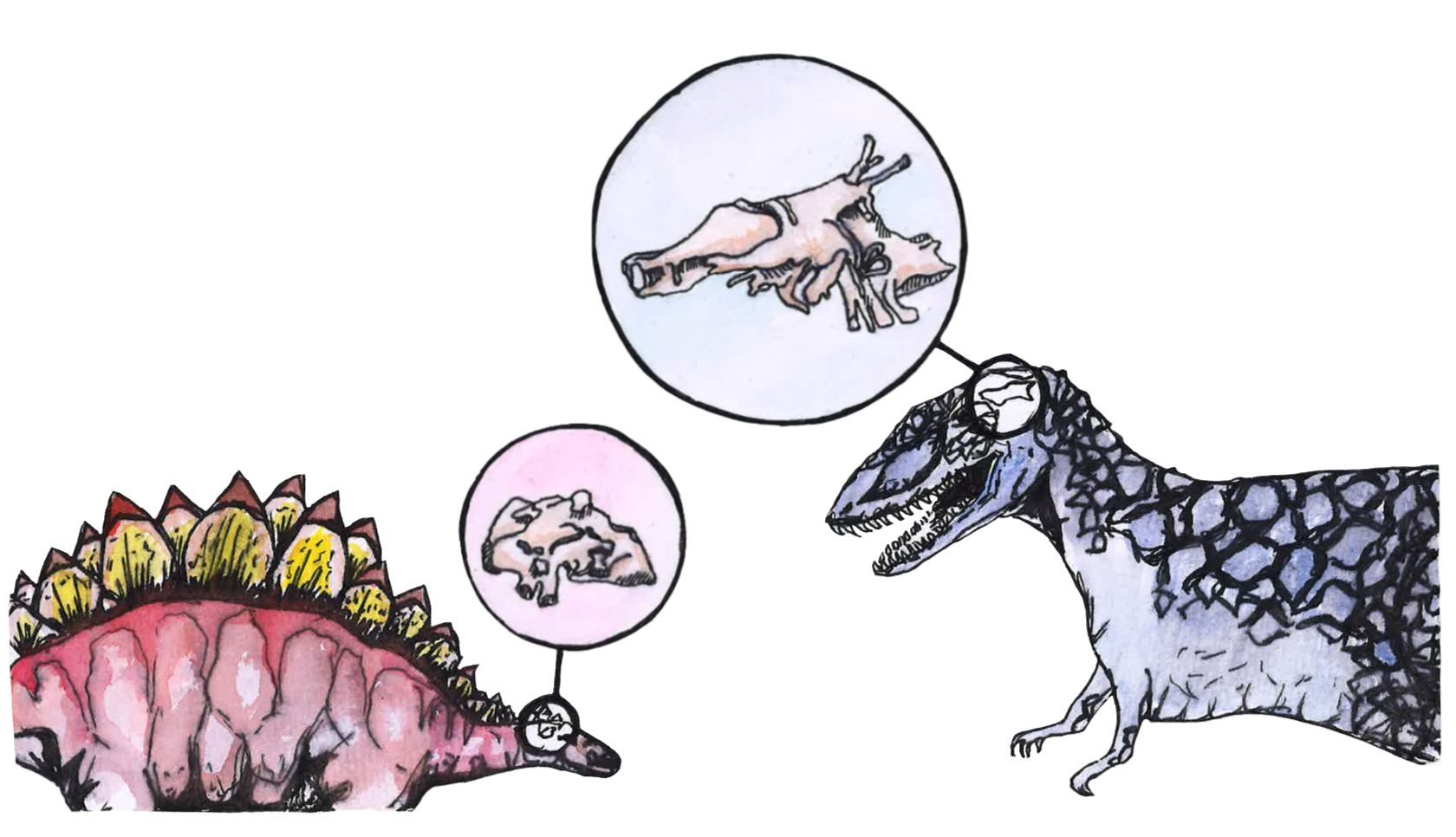
Before attempting to compare the brains of birds and dinosaurs, it is important to emphasize the diversity of dinosaurs across three primary groups: ornithischians, sauropods, and theropods [8]. Famous ornithischian dinosaurs include the three-horned Triceratops and the spiny Stegosaurus [17]. ‘Ornithischian’ in Greek can be translated to ‘bird-hipped,’ as ornithischians’ pelvic structures resemble those of modern-day birds, even though birds did not evolve from the ornithischian lineage [18]. Sauropods were similarly misnamed, since ‘sauropod’ means ‘lizard-footed’ in Greek, even though their gigantic feet actually bear little resemblance to those of modern-day lizards. The Brachiosaurus, Brontosaurus, and Titanosaurus make sauropods the lovable, gentle giants of the Jurassic Park franchise [1, 18, 19]. Finally, theropods are likely the most recognizable dinosaurs, including the famous Tyrannosaurus and Blue the Velociraptor, and are the most common subjects of paleoneurological study [8, 20]. Translating to ‘beast-footed’ in Greek, theropods primarily walked on two feet, ate mostly meat, had three toes or claws on each of their four limbs, and many were likely covered in feathers, much like birds [21]. Fittingly, modern-day birds evolved directly from theropods [7, 17].

With at least 66 million years of evolution differentiating them, modern-day birds and dinosaurs are functionally and anatomically distinct from one another [7, 14]. Still, birds share some key similarities with Avialae dinosaurs — the theropods most closely related to birds — that make them apt comparisons [21]. For example, the fossil record suggests that late avialan dinosaurs like Archaeopteryx and Anchiornis likely had wing and tail feathers, also implying the presence of the associated neurological mechanisms [10, 21]. Birds’ brain activity while completing cognitive tasks like solving puzzles can also be used to hypothesize which neurological capabilities may have been shared by dinosaurs with similar morphological characteristics [10]. Another avenue of inquiry comes from dinosaurs’ and birds’ next closest living relatives: crocodilians, a group of reptiles that includes alligators and crocodiles [17]. Crocodilians share a range of morphological similarities with many dinosaurs, including their large, pointed teeth and scaly exteriors [22]. Since birds and crocodilians are still alive, the features they share with dinosaurs provide clues about the behavior, lifestyles, and cognitive abilities of these extinct beasts [17].
Fossils: You Dig It?
Picture the classic image of a brain: a gelatinous, pinkish-gray mass. A soft and fleshy organ, the brain does not fossilize — only hard tissues like bone do [17]. So, when we use the fossil record to approximate dinosaur morphology and anatomy, we are limited to biological information that does not include the brain itself [10]. Using what evidence we do have, such as tooth shape or fossilized tracks, we can elucidate aspects of dinosaur behavior, including the general diet of a specific species, nesting sites and mating behavior, and preferred climate [22, 23]. For example, analysis of a site in Colorado suggests that theropods engaged in ceremonial courtship rituals similar to those of modern-day birds. This implies that theropods may have exhibited such flamboyant bird-like behaviors as flapping, dancing, and vocalizing [23, 24]. In the absence of brain tissue, paleoneurologists make cognitive inferences using fragments of dinosaur skulls, called endocasts, which are fossilized cranial cavities with indentations left by the brain [10]. Endocasts can provide neuroanatomical reference points that indicate the relative size of both the brain and its specialized regions using computed tomography (CT) scanning to digitally reconstruct the brain [17, 25, 26]. Imagine a T. rex being shuttled into a doughnut-shaped CT scanner — that’s what this technique attempts to simulate! Endocasts can reveal the relative sizes of the olfactory bulb, responsible for processing smell, or the flocculus, which plays a role in stabilizing vision and is generally larger in flying creatures than in terrestrial animals [27, 28, 29]. A larger olfactory bulb may indicate a well-developed sense of smell, an important trait for crocodilian hunting strategies [30]. Meanwhile, a larger flocculus points to greater agility or even flight, as in modern-day birds [27].
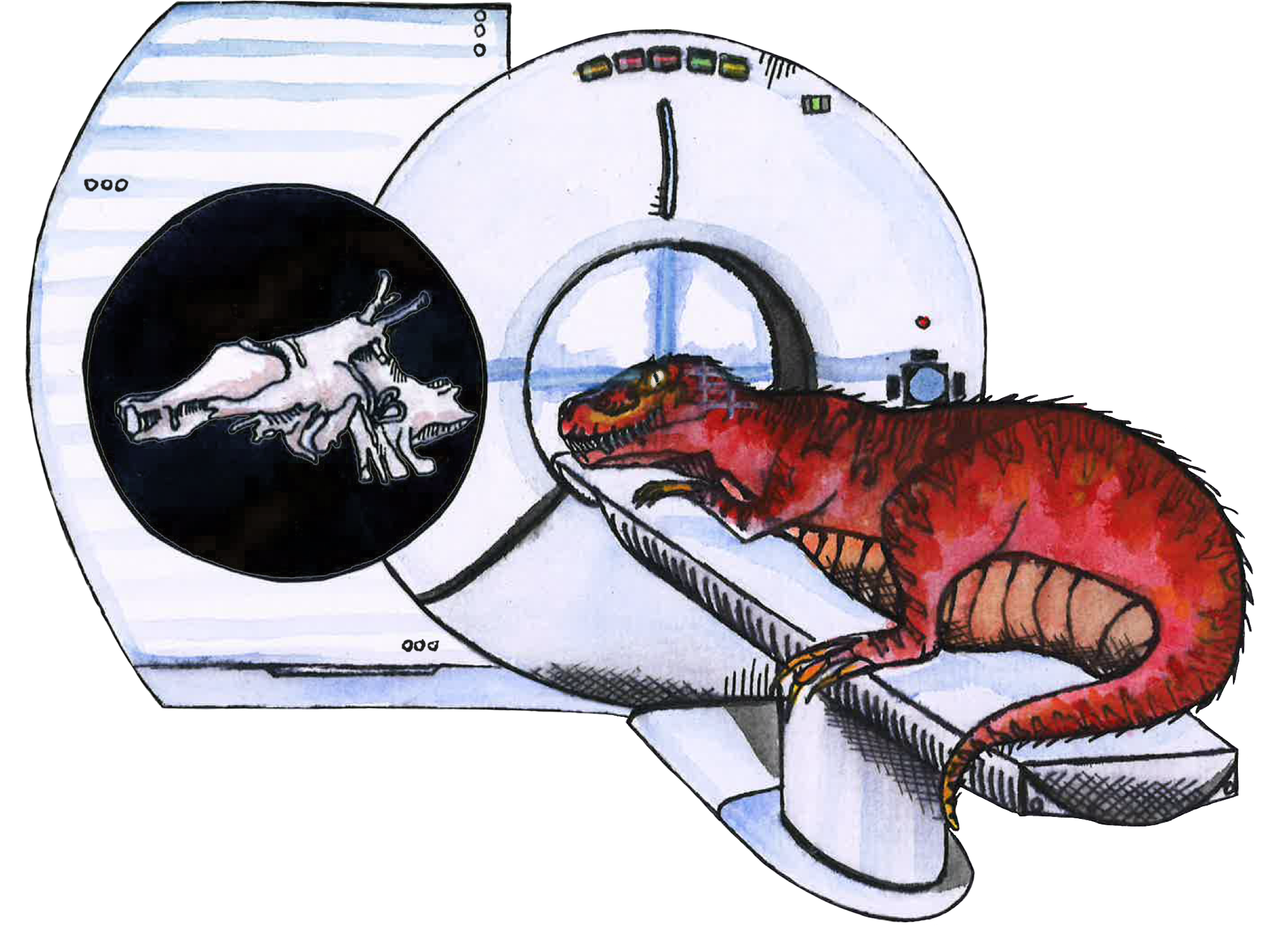
Endocasts are extremely useful for revealing sensory or anatomical traits, but discerning ‘intelligent’-type behaviors from endocasts is more challenging [31]. Indeed, it is difficult to identify levels of intelligence even in living animals, as defining what qualifies as an intelligent behavior or trait is in itself, quite a challenge [32, 33]. Debates surrounding the emotional intelligence, tool-use capacity, and social dynamics of, for instance, great apes — humans’ closest living relatives — have stumped comparative biologists for decades [34]. Tool use is widely regarded as a measure of intelligence, a trait once believed to singularly occur in humans, but has since been observed in various animals, including birds and crocodilians [32, 35, 36]. An additional marker of intelligence is problem-solving skills; innovation is associated with reduced extinction risk and larger brain size [37]. These metrics of intelligence can be associated with physical traits that endocasts and fossil evidence might reveal, such as opposable digits, a large brain cavity, bipedalism, and language centers [8]. Ultimately, intelligence is largely immeasurable and subjective, especially when applied to an extinct species [38]. Nevertheless, endocasts and fossil evidence can help uncover pieces of a larger cognitive puzzle [10].
The most straightforward method of endocast analysis is to assess total brain size [3]. An early hypothesis posits that brain size directly correlates with cognitive ability [33]. However, if this were true, whales, which have the largest recorded brains, would be the most intelligent animals on the planet [39]. Alternatively, relative brain size — the comparative size of an animal’s brain to its body — has a stronger correlation with intelligence and is quantified by the Encephalization Quotient (EQ) [3, 40]. EQ tends to be mammalian-centric, so other metrics have emerged for birds (BEQ) and reptiles (REQ) to more accurately compare EQ across species [36, 40]. Large theropods like T. rex tend to have normal-to-high BEQs and REQs comparable to those of living birds and reptiles, suggesting that theropods could have been as intelligent, or perhaps even more intelligent, than modern-day birds and reptiles [8, 36].
Another way to interpret endocasts is to examine the relative size of specific brain regions [41]. The mosaic brain evolution hypothesis posits that the brain expands unequally as it evolves, prioritizing brain real estate for specialized areas that are more relevant for a species’ specific functional needs [41, 42]. Consider the olfactory bulb in mice and humans: mice rely heavily on smell and therefore have proportionally larger olfactory bulbs than humans, who depend less on smell [43]. Similarly, comparing the relative sizes of brain regions from dinosaur endocasts with those of birds and crocodilians can reveal functional differences that point to lifestyle and cognitive traits in dinosaurs [27]. The boundaries between brain regions can be roughly identified by surface details left on the endocast that indicate the peaks and valleys of the outer layer of the brain [31]. The volumes of these brain regions can also be calculated with CT scanning and compared across species [31]. Ultimately, a broad analysis of all available data — fossil evidence, evolutionary relationships, and endocasts — can help reveal a comprehensive picture of dinosaurs’ lives [8]. Only then can we start to discern if they were brilliant strategic animals, brainless hunting machines, or somewhere in between [8].
Birdbrained: Or Were They?
Most dinosaur brains likely resembled bird brains, and no, this is not an insult [3, 8, 10]. Birds have wonderfully intricate brains that support many cognitive functions, such as complex vocal language — a trait otherwise only discovered in mammals — robust memory, innovative problem-solving, and tool use [32, 44, 45]. You are probably most familiar with the mammalian picture of a brain, with its wrinkly grayish-pink exterior. However, the mammalian brain has had over 300 million years to differentiate itself from the avian-reptilian brain, making its anatomy and processing mechanisms rather different [46, 47, 48]. Its wrinkly outer layer is the neocortex, the most cellularly dense region of the mammalian brain, which houses the most complex cognitive functions in mammals, including sensory integration, voluntary motor functions, and conscious mental processes [49]. Instead of a neocortex, birds and reptiles have a functionally equivalent structure called a pallium [50]. A major difference between the pallium and the neocortex lies in the optimization of surface area and volume [50]. The wrinkles of the neocortex allow for greater cellular connectivity, whereas the smoother avian-reptilian pallium hypothetically limits functional connectivity [8]. Because birds, reptiles, and dinosaurs all share a common ancestor, it is almost certain that dinosaur brains followed their pallial level of organization — a conclusion reinforced by endocast analysis [8, 47]. The avian pallium has an important substructure, the nidopallium, which is the seat of sophisticated decision-making in birds and has been linked to emotional, motivational, and perceptual processes [50, 51]. Since reptiles lack a nidopallium, evolutionary evidence alone cannot tell us whether dinosaurs had one [52]. Some dinosaur endocasts, however, reveal physical markings consistent with the presence of a nidopallium, though smaller in volume than those of modern-day birds [53]. If the presence of the nidopallium were confirmed, we could further infer the cognitive capabilities of dinosaurs, such as memory, decision-making, and innovation [53].
Collectively, evolutionary and endocast evidence support the argument that dinosaurs’ brains were both organizationally and functionally comparable to those of birds [3, 8]. Therefore, we can speculate that dinosaurs may have possessed some of the sophisticated cognitive abilities of birds [3]. For instance, many bird species have complex vocal language that rivals humans’ and may even use grammatical structure [44]. It is possible that dinosaurs were capable of vocalizations similar to those made by modern-day birds — a theory supported by fossil evidence of a similar voice box in both birds and dinosaurs [54]. Vocal learning in birds has been associated with larger brain size and greater problem-solving abilities, so bird-like vocalizations in dinosaurs may indicate enhanced cognitive abilities [37]. Many birds, such as crows, have demonstrated robust memories, innovation, and tool use [45]. Likewise, it has been speculated that dinosaurs could have used tools, based on what fossil evidence suggests about the dexterity of their limbs and digits [36]. Essentially, dinosaurs were likely not as cognitively complex as primates, as some have suggested, solely because of the organization of their brains [3, 8]. However, birds — which represent in brain structure what dinosaur brains probably looked like — are very intelligent animals [37]. So, dinosaurs, too, could have been as intelligent as their feathered descendants, except much, much toothier [8].
Cul-de-Sac Cognition
In addition to differences in the structural organization of mammalian and avian brains, they are also distinct in their organization at the cellular level [8, 50, 55]. The mammalian neocortex is highly organized and densely packed with neurons, or brain cells, that form six distinct layers and many functional columns, like an apartment building with hallways connecting units and an elevator between floors [56]. In contrast, the avian-reptilian pallium is more nuclear and radial in its neural organization, forming neural clusters, like a neighborhood with numerous cul-de-sacs off which houses radiate [52]. You can see how it might be easier to deliver a letter from one apartment to another than to drive all the way from one house on a cul-de-sac to a second house on a different cul-de-sac. The cul-de-sac model of organization theoretically limits the extent to which the avian-reptilian brain can expand and adapt, especially compared to the efficiency of the human brain [8, 52]. For instance, neuroplasticity — the capacity for change between connections in the brain — is a key process behind learning (57). In a nuclear system, neuroplasticity may be restricted because neurons are less connected to one another than in the layers of the mammalian neocortex [8]. The means of communication between neurons, like the houses, are restricted to travel along a central road, whereas in the apartment building, you could yell your message through the vents to reach the floor above you or climb the fire escape. Essentially, mammalian layered organization theoretically provides more avenues for connection and speed of transmission. While we can only hypothesize about the consequences of pallial organization on dinosaur intelligence, one theorist has used a comparative approach towards endocasts to defend the notion that dinosaurs could not have reached mammalian levels of sophistication [8]. However, these conclusions are based on research that is focused predominantly on mammalian learning and brains rather than on birds — an example of mammalian-centric bias in cognitive research [50, 55].
The vast size difference between dinosaurs and birds makes it even harder to know the degree to which the neural organization of bird brains can actually apply to dinosaur brains [3, 8]. The effect of scaling up bird brains to dinosaur proportions would impact both neuron density and neural length; in some cases, neural density is even more indicative of intelligence than EQ [8, 58]. Based on a generous scaling technique, one estimate suggests that dinosaurs’ neuron abundance was equal to that of primates, a conclusion that is highly debated [3, 17]. A separate endocast technique assumes that the T. rex cranial cavity was occupied less by brain tissue itself and more by fluid, characteristically more like crocodilians and less like birds. This endocast approach yields a lower brain tissue volume estimate and suggests that T. rex’s neural density was more similar to that of crocodilians than primates [17]. Together, these estimates highlight both the importance of evolutionary relationships and the challenges of relying on them to make inferences [10].
Fossil Fools: The Limitations of Paleoneurology
The tools that have been developed to piece together dinosaur cognitive abilities, namely endocasts and evolutionary comparisons, are helpful but imperfect, as they are subject to human bias and interpretation [5, 10]. A trend in comparative biology is the tendency to privilege our own brand of human intelligence, a line of reasoning that often places humans at the top of an evolutionary pyramid [59, 60]. The qualities and behaviors used to define intelligence, such as tool use, language, or problem-solving, are classic benchmarks of human cognition, and quantifying these traits in nonhuman animals may not be realistic [5, 8]. Take vision, for example. Humans’ vision is restricted to a narrow band of light on the electromagnetic spectrum we call ‘visible light’ — a term that reflects our anthropocentrism [61, 62]. However, many birds’ eyes are equipped to see not only visible light but also ultraviolet (UV) light, allowing them to detect natural patterns on prey, food, and mates that we cannot [61]. Based on eye structure evidence, it is possible that some bird-like theropods were also capable of UV vision [63]. These dinosaurs would have seen the Earth in quite literally a whole different light, illustrating how narrow our experiences are when compared across the animal kingdom [63]. To bridge this knowledge gap, some researchers have proposed a context-dependent approach that frames cognitive success as a species’ ability to adapt and thrive under its unique environmental pressures [5]. In this sense, T. rex’s tiny arms, razor-sharp teeth, and bloodhound-like nose reflect its brain’s ability to coordinate the motor and hunting skills required to become an apex predator in its ecological niche [64, 65, 66]. To understand how intelligent dinosaurs were, we must first recognize that being a dinosaur meant having an entirely different brain, body, and environment than our own [5].
The Saur Losers
The evidence for dinosaur intelligence remains subject to debate, and we may never know how these magnificent creatures ruled their version of Earth [8, 10, 17]. Still, we can respect the longevity of their mighty 160-million-year reign [6]. While humans may possess more ‘intelligence’ than our reptilian predecessors, we have inflicted permanent damage onto the planet in just 300,000 years: one million animal and plant species are currently threatened with extinction, and countless terrestrial and marine environments have been significantly altered over the course of humans’ reign [67, 68]. The unprecedented destruction of the human era may be the very reason that the ‘dinosaur age’ seems so idyllic [4]. However, modern technology may be able to repair some human-inflicted harm, with the new possibility of resurrecting extinct species like the woolly mammoth, the passenger pigeon, and, yes, the dinosaurs [69]. The existence of a real Jurassic Park may not be such a far-off fantasy, but would this reality be as picturesque as the screen portrays? The beloved franchise poses the question: Do we have the right to interfere with the laws of nature, and what are the potential pitfalls of our intelligence? Perhaps human-like intelligence is not to be coveted at all, but rather something that will destroy the world as we know it, like a self-made asteroid strike.
References
Spielberg, S. (Director). (1993). Jurassic Park [Film]. Universal Pictures; Amblin Entertainment.
Bayona, J.A. (Director). (2018). Jurassic World: Fallen Kingdom [Film]. Universal Pictures; Amblin Entertainment; Legendary Pictures; The Kennedy/Marshal Company; Perfect World Pictures.
Herculano-Houzel, S. (2023). Theropod dinosaurs had primate-like numbers of telencephalic neurons. Journal of Comparative Neurology, 531(9), 962–974. doi: 10.1002/cne.25453
Reiner, A. (2023). How smart could dinosaurs get? The Journal of Comparative Neurology, 531(9), 956–958. doi: 10.1002/cne.25471
Jensen, T. R., Jacobs, I., Kverková, K., Lalić, L., Polonyiová, A., Stehlík, P., Reber, S. A., & Osvath, M. (2025). T. rex cognition was T. rex-like—A critical outlook on diverging views of the neurocognitive evolution in dinosaurs. The Anatomical Record. doi: 10.1002/ar.70074
Chiarenza, A. A. (2024). The macroecology of Mesozoic dinosaurs. Biology Letters, 20(11), 20240392. doi: 10.1098/rsbl.2024.0392
Benson, R. B. J. (2018). Dinosaur macroevolution and macroecology. Annual Review of Ecology, Evolution, and Systematics, 49(Volume 49), 379–408. doi: 10.1146/annurev-ecolsys-110617-062231
Reiner, A. (2023). Could theropod dinosaurs have evolved to a human level of intelligence? Journal of Comparative Neurology, 531(9), 975–1006. doi: 10.1002/cne.25458
Thibault, S., Py, R., Gervasi, A. M., Salemme, R., Koun, E., Lövden, M., Boulenger, V., Roy, A. C., & Brozzoli, C. (2021). Tool use and language share syntactic processes and neural patterns in the basal ganglia. Science, 374(6569), eabe0874. doi: 10.1126/science.abe0874
Balanoff, A. M. (2024). Dinosaur palaeoneurology: An evolving science. Biology Letters, 20(12), 20240472. doi: 10.1098/rsbl.2024.0472
Mannion, P. D. (2024). The spatiotemporal distribution of Mesozoic dinosaur diversity. Biology Letters, 20(12), 20240443. doi: 10.1098/rsbl.2024.0443
Chiarenza, A. A., Farnsworth, A., Mannion, P. D., Lunt, D. J., Valdes, P. J., Morgan, J. V., & Allison, P. A. (2020). Asteroid impact, not volcanism, caused the end-Cretaceous dinosaur extinction. Proceedings of the National Academy of Sciences, 117(29), 17084–17093. doi: 10.1073/pnas.2006087117
Condamine, F. L., Guinot, G., Benton, M. J., & Currie, P. J. (2021). Dinosaur biodiversity declined well before the asteroid impact, influenced by ecological and environmental pressures. Nature Communications, 12. doi: 10.1038/s41467-021-23754-0
Brusatte, S. L., O’Connor, J. K., & Jarvis, E. D. (2015). The origin and diversification of birds. Current Biology, 25(19), R888–R898. doi: 10.1016/j.cub.2015.08.003
James, F. C. (2021). How many dinosaurs are birds? BioScience, 71(9), 991–994. doi:10.1093/biosci/biab060
Field, D. J., Burton, M. G., Benito, J., Plateau, O., & Navalón, G. (2025). Whence the birds: 200 years of dinosaurs, avian antecedents. Biology Letters, 21(1), 20240500. doi: 10.1098/rsbl.2024.0500
Caspar, K. R., Gutiérrez-Ibáñez, C., Bertrand, O. C., Carr, T., Colbourne, J. A. D., Erb, A., George, H., Holtz Jr, T. R., Naish, D., Wylie, D. R., & Hurlburt, G. R. (2024). How smart was T. rex? Testing claims of exceptional cognition in dinosaurs and the application of neuron count estimates in palaeontological research. The Anatomical Record, 307, 3685–3716. doi: 10.1002/ar.25459
Sander, P. M. (2023). Sauropods. Current Biology, 33(2), R52–R58. doi:10.1016/j.cub.2022.12.024
Edwards, G. (Director). (2025). Jurassic World Rebirth [Film]. Universal Pictures; Amblin Entertainment.
Rowe, A. J., & Rayfield, E. J. (2025). Carnivorous dinosaur lineages adopt different skull performances at gigantic size. Current Biology, 35(15), 3664–3673. doi: 10.1016/j.cub.2025.06.051
Agnolin, F. L., Motta, M. J., Brissón Egli, F., Lo Coco, G., & Novas, F. E. (2019). Paravian phylogeny and the dinosaur-bird transition: An overview. Frontiers in Earth Science. doi: 10.3389/feart.2018.00252
Kundanati, L., D’Incau, M., Bernardi, M., Scardi, P., & Pugno, N. M. (2019). A comparative study of the mechanical properties of a dinosaur and crocodile fossil teeth. Journal of the Mechanical Behavior of Biomedical Materials, 97, 365–374. doi: 10.1016/j.jmbbm.2019.05.025
Lockley, M. G., McCrea, R. T., Buckley, L. G., Deock Lim, J., Matthews, N. A., Breithaupt, B. H., Houck, K. J., Gierliński, G. D., Surmik, D., Soo Kim, K., Xing, L., Yong Kong, D., Cart, K., Martin, J., & Hadden, G. (2016). Theropod courtship: Large scale physical evidence of display arenas and avian-like scrape ceremony behaviour by Cretaceous dinosaurs. Scientific Reports, 6. doi: 10.1038/srep18952
Soma, M., & Iwama, M. (2017). Mating success follows duet dancing in the Java sparrow. PLOS ONE, 12(3), e0172655. doi: 10.1371/journal.pone.0172655
Hu, J., Xu, X., Zhao, Q., He, Y., Forster, C. A., & Han, F. (2024). Endocranial morphology of three early-diverging ceratopsians and implications for the behavior and the evolution of the endocast in ceratopsians. Paleobiology, 50(3), 490–502. doi: 10.1017/pab.2024.25
Bruner, E. (2018). The brain, the braincase, and the morphospace. In E. Bruner, N. Ogihara, & H. Tanabe (Eds.), Digital Endocasts: From Skulls to Brains (pp. 93–114). Springer Japan. doi: 10.1007/978-4-431-56582-6_7
Belén von Baczko, M., Cardillo, A. F., Ulloa-Guaiquin, K., Desojo, J. B., & Paulina-Carabajal, A. (2024). Paleoneurology and neuroanatomical notes on the South American “rauisuchian” Saurosuchus galilei from the Ischigualasto formation, San Juan, Argentina. The Anatomical Record, 307, 1239–1253. doi: 10.1002/ar.25362
Paulina-Carabajal, A., & Currie, P. J. (2017). The braincase of the theropod dinosaur Murusraptor: Osteology, neuroanatomy and comments on the paleobiological implications of certain endocranial features. Ameghiniana, 54(5), 617–640. doi: 10.5710/AMGH.25.03.2017.3062
Zaidi, S., Yasir, H., & Jaka, A. (2024). Floccular syndrome—An atypical presentation of paraneoplastic cerebellar degeneration. Journal of Ayub Medical College Abbottabad, 36(3). doi: 10.55519/JAMC-03-12493
Dumont Jr, M. V., Santucci, R. M., de Andrade, M. B., & de Oliveira, C. E. M. (2020). Paleoneurology of Baurusuchus (Crocodyliformes: Baurusuchidae), ontogenetic variation, brain size, and sensorial implications. The Anatomical Record, 305(10), 2670–2694. doi: 10.1002/ar.24567
Labra, N., Mounier, A., Leprince, Y., Rivière, D., Didier, M., Bardinet, E., Santin, M. D., Mangin, J. F., Filippo, A., Albessard-Ball, L., Beaudet, A., Broadfield, D., Bruner, E., Carlson, K. J., Cofran, Z., Falk, D., Gilissen, E., Gómez-Robles, A., Neubauer, S., … Balzeau, A. (2024). What do brain endocasts tell us? A comparative analysis of the accuracy of sulcal identification by experts and perspectives in palaeoanthropology. Journal of Anatomy, 244(2), 274–296. doi: 10.1111/joa.13966
Kabadayi, C., & Osvath, M. (2017). Ravens parallel great apes in flexible planning for tool-use and bartering. Science, 357(6347), 202–204. doi: 10.1126/science.aam8138
van Schaik, C. P., Triki, Z., Bshary, R., & Heldstab, S. A. (2021). A farewell to the encephalization quotient: A new brain size measure for comparative primate cognition. Brain, Behavior and Evolution, 96(1), 1–12. doi: 10.1159/000517013
Leavens, D. A., Bard, K. A., & Hopkins, W. D. (2019). The mismeasure of ape social cognition. Animal Cognition, 22, 487–504. doi: 10.1007/s10071-017-1119-1
Wink, M. (2022). Intelligence in the animal kingdom. In R. M. Holm-Hadulla, J. Funke, & M. Wink (Eds.), Intelligence—Theories and Applications (pp. 87–103). Springer. doi: 10.1007/978-3-031-04198-3_4
Varricchio, D., Hogan, J., & Freimuth, W. (2021). Revisiting Russell’s troodontid: Autecology, physiology, and speculative tool use. Canadian Journal of Earth Sciences. doi: 10.1139/cjes-2020-0184
Audet, J.-N., Couture, M., Lefebvre, L., & Jarvis, E. D. (2024). Problem-solving skills are predicted by technical innovations in the wild and brain size in passerines. Nature Ecology & Evolution, 8, 806–816. doi: 10.1038/s41559-024-02342-7
Glock, H.-J. (2019). Agency, intelligence and reasons in animals. Philosophy, 94(4), 645–671. doi: 10.1017/S0031819119000275
Burger, J. R., George Jr., M. A., Leadbetter, C., & Shaikh, F. (2019). The allometry of brain size in mammals. Journal of Mammalogy, 100(2), 276–283. doi: 10.1093/jmammal/gyz043
Lauters, P., Vercauteren, M., & Godefroit, P. (2023). Endocasts of ornithopod dinosaurs: Comparative anatomy. In Progress in Brain Research (Vol. 275, pp. 1–23). Elsevier. doi: 10.1016/bs.pbr.2022.10.002
Felice, R. N., & Goswami, A. (2018). Developmental origins of mosaic evolution in the avian cranium. Proceedings of the National Academy of Sciences, 115(3), 555–560 doi: 10.1073/pnas.1716437115
Fong, S., Rogell, B., Amcoff, M., Kotrschal, A., van der Bijl, W., Buechel, S. D., & Kolm, N. (2021). Rapid mosaic brain evolution under artificial selection for relative telencephalon size in the guppy (Poecilia reticulata). Science Advances, 7(46). doi: 10.1126/sciadv.abj4314
McGann, J. P. (2017). Poor human olfaction is a 19th-century myth. Science, 356(6338), eaam7263. doi: 10.1126/science.aam7263
Spierings, M. J., & ten Cate, C. (2016). Budgerigars and zebra finches differ in how they generalize in an artificial grammar learning experiment. Proceedings of the National Academy of Sciences, 113(27), E3977–E3984. doi: 10.1073/pnas.1600483113
Balakhonov, D., & Rose, J. (2017). Crows rival monkeys in cognitive capacity. Scientific Reports, 7(1), 8809. doi: 10.1038/s41598-017-09400-0
Zou, Y., Zhang, Z., Zeng, Y., Hu, H., Hao, Y., Huang, S., & Li, B. (2024). Common methods for phylogenetic tree construction and their implementation in R. Bioengineering, 11(5), 480. doi: 10.3390/bioengineering11050480
Shimizu, T., Shinozuka, K., Uysal, A. K., & Leilani, S. (2017). The origins of the bird brain: Multiple pulses of cerebral expansion in evolution. In S. Watanabe, M. A. Hofman, & T. Shimizu (Eds.), Evolution of the Brain, Cognition, and Emotion in Vertebrates (pp. 35–57). Springer Japan. doi: 10.1007/978-4-431-56559-8_2
Ksepka, D. T., Balanoff, A. M., Smith, N. A., Bever, G. S., Bhullar, B.-A. S., Bourdon, E., Braun, E. L., Burleigh, J. G., Clarke, J. A., Colbert, M. W., Corfield, J. R., Degrange, F. J., De Pietri, V. L., Early, C. M., Field, D. J., Gignac, P. M., Gold, M. E. L., Kimball, R. T., Kawabe, S., Lefebvre, L., Marugán-Lobón, J., Mongle, C.S., Morhadt, A., Norell, M.A., Ridgely, R.C., Rothman, R.S., Scofield, R.P., Tambussi, C.P., Toress, C.R., van Tuinen, M., Walsh, S.A., Watanabe, A., Witmer, L.M., Wright, A.K., Zanno, L.E., Jarvis, E.D., Smaers, J. B. (2020). Tempo and pattern of avian brain size evolution. Current Biology, 30(11), 2026-2036.e3. doi: 10.1016/j.cub.2020.03.060
Kass, J. H., Qi, H.-X., Reed, J. L., & Krubitzer, L. A. (2026). The evolution of neocortex. In J. H. Kaas & S. Herculano-Houzel (Eds.), Evolution of Nervous Systems (3rd ed., Vol. 2, pp. 163–182). Academic Press.doi: 10.1016/B978-0-443-27380-3.00034-8
Güntürkün, O., von Eugen, K., Packheiser, J., & Pusch, R. (2021). Avian pallial circuits and cognition: A comparison to mammals. Current Opinion in Neurobiology, Evolution of Brains and Computation, 71, 29–36. doi: 10.1016/j.conb.2021.08.007
Carlén, M. (2017). What constitutes the prefrontal cortex? Science, 358(6362), 478–482. doi: 10.1126/science.aan8868
Puelles, L., Sandoval, J. E., Ayad, A., del Corral, R., Alonso, A., Ferran, J. L., & Martínez-de-la-Torre, M. (2017). The pallium in reptiles and birds in the light of the updated tetrapartite pallium model. In Evolution of Nervous Systems: Second Edition (Vol. 1, pp. 519–555). doi: 10.1016/B978-0-12-804042-3.00014-2
Gaetano, T. M., Yacobucci, M. M., & Bingman, V. P. (2017). On the paleontology of animal cognition: Using the brain dimensions of modern birds to characterize maniraptor cognition. Journal of Advanced Neuroscience Research, 12–19. doi: 10.15379/2409-3564.2017.03
Yoshida, J., Kobayashi, Y., & Norell, M. A. (2023). An ankylosaur larynx provides insights for bird-like vocalization in non-avian dinosaurs. Communications Biology, 6, 152. doi: 10.1038/s42003-023-04513-x
Rose, J. (2022). The avian brain. Current Biology, 32(20), R1076–R1079. doi: 10.1016/j.cub.2022.07.072
Hawkins, J., Lewis, M., Klukas, M., Purdy, S., & Ahmad, S. (2019). A framework for intelligence and cortical function based on grid cells in the neocortex. Frontiers in Neural Circuits, 12. doi: 10.3389/fncir.2018.00121
Mateos-Aparicio, P., & Rodríguez-Moreno, A. (2019). The impact of studying brain plasticity. Frontiers in Cellular Neuroscience, 13. doi: 10.3389/fncel.2019.00066
Olkowicz, S., Kocourek, M., Lučan, R. K., Porteš, M., Fitch, W. T., Herculano-Houzel, S., & Němec, P. (2016). Birds have primate-like numbers of neurons in the forebrain. Proceedings of the National Academy of Sciences, 113(26), 7255–7260. doi: 10.1073/pnas.1517131113
Font, E., Burghardt, G. M., & Leal, M. (2023). Brains, behaviour, and cognition: Multiple misconceptions. In C. Warwick, P. C. Arena, & G. M. Burghardt (Eds.), Health and Welfare of Captive Reptiles. Springer. doi: 10.1007/978-3-030-86012-7_7
Wiest, G. (2025). Hierarchical concepts in neuroethology. In Hierarchies in the Brain, Mind and Behaviour: A Principle Of Neural and Mental Function (pp. 31–93). Springer Nature Switzerland. doi: 10.1007/978-3-031-78030-1_3
Cronin, T. W., & Bok, M. J. (2016). Photoreception and vision in the ultraviolet. Journal of Experimental Biology, 219(18), 2790–2801. doi: 10.1242/jeb.128769
Kopnina, H., Washington, H., Taylor, B., & Piccolo, J. J. (2018). Anthropocentrism: More than just a misunderstood problem. Journal of Agricultural and Environmental Ethics, 31(1), 109–127. doi: 10.1007/s10806-018-9711-1
Woodruff, D. C., Naish, D., & Dunning, J. (2021). Photoluminescent visual displays: An additional function of integumentary structures in extinct archosaurs? Historical Biology, 33(9), 1718–1725. doi: 10.1080/08912963.2020.1731806
Gignac, P. M., & Erickson, G. M. (2017). The Biomechanics Behind Extreme Osteophagy in Tyrannosaurus rex. Scientific Reports, 7, 2012. doi: 10.1038/s41598-017-02161-w
Hughes, G. M., & Finarelli, J. A. (2019). Olfactory receptor repertoire size in dinosaurs. Proceedings of the Royal Society B: Biological Sciences, 286(1904). doi: 10.1098/rspb.2019.0909
Holtz Jr., T. R. (2021). Theropod guild structure and the tyrannosaurid niche assimilation hypothesis: Implications for predatory dinosaur macroecology and ontogeny in later Late Cretaceous Asiamerica. Canadian Journal of Earth Sciences, 58(9), 778–795. doi: 10.1139/cjes-2020-0174
Galway-Witham, J., & Stringer, C. (2018). How did Homo sapiens evolve? Science, 360(6395), 1296–1298. doi: 10.1126/science.aat6659
Turvey, S. T., & Crees, J. J. (2019). Extinction in the Anthropocene. Current Biology, 29(19), R982–R986. doi: 10.1016/j.cub.2019.07.040
Seddon, P. J. (2017). The ecology of de-extinction. Functional Ecology, 31(5), 992–995. doi: 10.1111/1365-2435.12856